Notes of 1st Semester (Core), Zoology Evolution of symmetry & segmentation of Metazoa.pdf - Study Material
Page 1 :
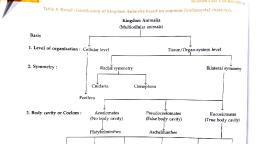
Sos Sreeens mee Te WO THITLOP NRE halved Are HOLsiperinposable and, thereby ave recognisable ax roxpectively the right and left half,, , Origin and Evolution of Symmotry, , Understanding, the evolutionary origin of novel me, of the most souphtaler answers in biology. P, occurred in the Metazoa is, , lazoun body plans continues to be one, erhaps the most profound change that may have, the appearance Of bilaterally symmetrical forms from a presumably, tadially symmetrical ancestor, ‘The symmetry properties of bilaterally symmetrical larval, and adult metazouns are generally set up during, the cleavage period while most “radially”, symmetrical cnidarians do not display a stereotyped cleavage program, Ctenophores display, biradial symmetry and may represent one intermediate form in the transition to bilateral, symmetry, The origin of the dorsal-ventral axis, and indeed the relationships of the major, longitudinal axes, in enidarians, clenophores, and bilaterian animals are however far from, certain, The realization that many of the molecular mechanisms of axial determination are, conserved throughout the Bilateria allows one to formulate a set of predictions as to their, possible role in the origins of bilateria. ‘The basic ideas prevailing in biology related to the, evolution of symmetry can be found in the theories on the origin of metazoa, which are, discussed as follows,, , I. Syncytial Ciliate Theory: Syncytial theory of origin of metazoans was first proposed, by Jovan Hadzi (1953) and by Earl D. Hanson (1958). Their proposition was based on, resemblance between multinucleate ciliates and acoclomate flatworms. Syncytial refers to the, multinucleate condition in which cell membranes are absent between adjacent nuclei. This, theory is called syncytial theory (Fig. 10.3) because it emphasises that metazoans (acoclomate, flatworm) have evolved from the segmentation of multinucleate ciliate, Later, cell membrane, developed around the nuclei and different cells differentiated structurally and functionally, It is, assumed that the body form of the ancestor resembled that of modern ciliates and thus tended, toward bilateral symmetry. Therefore, the earliest metazoans would have been bilateral and, similar to some extant flatworms., , , , Scanned with CamScanner
Page 2 :
Multinucleated Acoelomate metazoon, ciliated protist precursor after, Syncytical cell cellularization, , , , Fig. 10.3. Syncytical theory, , , , , , , , There are several objections to this hypothesis. It ignores embryology of the flatworms in, which nothing similar to cellularization occurs; it does not explain the presence of flagellated, sperm in metazoans; and, perhaps more important, it implies that the radial symmetry of, cnidarians is derived from a primary bilateral symmetry., , 2. Colonial Flagellate Theory: The colonial flagellate theory, first proposed by Haeckel, in 1874, is the classical scheme, which, with various revisions, still has many followers. The, theory was first proposed by Emst Haeckel (1874) and an offshoot theory was proposed by, Otto Butschli (1883). It was modified by Metschnikoff (1887). This theory was later revived, by L.H. Hyman (1940). There are many variations of this theory:, , (a) Haeckel’s Theory of Gastrea: The colonial theory is mainly based on proposition, made in Haeckel’s (1874) Gasterea Theory of metazoan ancestory (Fig. 10.4 a). Metazoan, evolved by aggregation of single protistan cells into little hollow, spherical, flagellate colony,, similar to Volvox with distinct antero-posterior axis. Individual cells within the colony became, differentiated for specific functional roles (reproductive cells, nerve cells, somatic cells, and, so on), thus subordinating cellular independence to welfare of the colony as a whole. The, colonial ancestral form was at first radially symmetrical and reminiscent of a blastula stage of, development. This hypothetical ancestor was called a blastaea. Drawing on the developmental, sequence of extant animals as a model, Haeckel hypothesized that ancestral forms similar to a, gastrula may have existed. These ancestors were called gastraea. Cnidarians, with their radial, symmetry, could have evolved from this form., , The main criticism to this theory is that endoderm is not always formed by invagination but, it can be formed by inwandering of the ectodermal cells. Also, in cnidarians gastrulas are solid, not hollow. Again, according to Haeckel, radial symmetry is regarded as primitive character, whereas it is also seen in case of echinoderms and some lower chordates where the larval, form is bilaterally symmetrical and adults are radially symmetrical (adaptation to sedentary, lifestyle)., , Scanned with CamScanner
Page 3 :
Solid Gastrea, , , , Fig. 10.4. Two versions of colonial flagellate theory, (a) Haeckel’s theory (b)Metschnikoff’s theory, , , , , , , , (6) Metschnikoff’s Revised Theory of Gastrea: The Theory of Gastrea by Haeckel (1874), was modified by Metchnikoff (1887). According to Metchnikoff’s observation the primitive, mode of gastrulation in cnidarian is by ingression (inwandering of ectodermal cells) and not by, invagination as suggested by Haeckel’s theory. So, he suggested that blastea didn’t invaginate,, it underwent ingression. He also discovered that ingression resulted in the solid gastrula, characteristic of cnidarians. The ancestor was called Planuloid and the theory was called, The Planuloid Theory. Planuloid was a radially symmetrical ovoid, solid mass of cells with, exterior flagella (Fig 10.4 b). There was no mouth and exterior phagocytosis brought in the, food. This was similar to the free-swimming planula larva of coelenterates and was considered, as a prospective ancestor to all other metazoans. It explains the cnidarian radial symmetry., , The major drawback to this proposition is that no living representatives other than the, planuloid life stage of cnidaria has been reported. :, , (c) Otto Butschli’s Theory of Plakula: The Theory of Plakula was proposed by Butschli, (1883). According to Otto Butschli (1883) no model could explain the purpose of evolution of, a digestive layer which is more likely to be associated with absorption than external layer. The, Plakula theory states that, most metazoans after sponges were flattened disc shaped mass of cells, with two layers of flagellated cells — the ventral being digestive and dorsal being protective and, reproductive cells. They were benthic organisms which creep on substrate and ventral surface, is modified for absorption and would hunch up to capture, digest food. This stage would have, given rise to other forms with more permanent cavity. The proposed progression was from, creeping existence to bilateral symmetry and finally to bilaterogastrea., , This theory too has been criticized on accounts that it does not explain radial symmetry in, , Cnidaria., a, , Scanned with CamScanner
Page 4 :
Se, , RO ET, (dO) Hymen\ Colonial Theory: Hynven (1940) supported hollow bhasties ehow of Haeekel,, , but added that due to multipolar ingression, hollow blistwea became salid and eisdially, , symmetrical Parenchymula stereogastrula of Metechniiotl CN) looming planulotd rivtacean, , ancestor (similar to planula larva of coelenterates), , A. Polyphyletic Theory of Metazoan Originy This theory ermpliasioes on origin ard, evolution of different group of ingdom Animalia ta polyphyletic, (e Matwrortes (plararions), , arose due to cellulanzation of syncytial multinucleate unicellular ciliate protovoans ane the, sponges arose separately trom choanoflagelates, , There are thus several ideas concerning the origin of bilateral symmetry within the, Metazoa. If one accepts Hadzi's syncytial theory or Butschli's plakula, then the problent ps, already solved, since bilaterality would characterize the first Metazoa. On the other hand,, supporters of the colonial theory generally assume that the first bilateral animals arose fron, a gastrula-like ancestor, presumably one with spherical or radial symmetry, of ite planulotd, descendant, It has been suggested that the blastea took up a benthic lifestyle, invaginated, and, , then assumed bilaterality, In this scenario, the first metazoan was a bilaterally symmetrical, gastrea (bilaterogastrea) from which all other major groups arose, , METAMERISM/SEGMENTATION, , , , oo, , Metamerism is the serial repetition of similar body sections along the longitudinal axis of, the body with each part containing similar components of all major organ systems. Bach such, section or part is called a segment, somite or metamere. The terms metamerism or metameric, segmentation are used only when organs of mesodermal origin are so arranged, Hach tmetamere, typically repeats some or all of the various organ units. The primary segmental divisions are, the body wall musculature and sometimes the coelom. This in turn imposed a corresponding, Meltamerism on the associated supply systems—nerves, blood vessels and excretory organs, In, other words, all parts which do not run the length of the animal are serially repeated at regular, intervals along the axis, and it is this repetition which constitutes the essence of metameriam., Metamerism is always limited to the trunk region of the body. The head, represented by the, Prostomium and: bearing the brain and sense organs, and the pygidium, represented by the, terminal part of the body which carries the anus, are not metameres, Evolutionary changes have, ‘oday obscured much of the segmentation in many animals, including humans., , Occurrence of Metamerism in Animal Kingdom, , All members of three large animal phyla are metameric: Annelida, Arthropoda, and, Ordata. The first two exhibit conspicuous segmentation in the adult. Among the chordates,, c eu metameric pattern is evident in muscles, vertebrae, and ribs of the adult (e.g,, Shes), but even when less obvious (c.g. mammals), the development of each individual is, Sed firmly on formation of segments, t the embryological somites. Segments of the tapeworm, , Scanned with CamScanner
Page 5 :
es NON-CHORDATES: PROTISTS TO PSEUDOCOELOMaTES | te, , (proglottids) are formed so differently from the segments of the other three groups that most!, zoologists do not admit tapeworms to be metamerically segmented animals. Metamerism!, of Annelida and Arthropoda and that of Chordata probably arose independently and hence |, metamerism does not itself imply relationships between the groups however, the particular, , metamerism within each group clearly demonstrates the derivative relationship of its members,, , Types of Metamerism, , Following types of metamerism are generally distinguished within the animal kingdom,, , 1. Homonomous and Heteronomous Metamerism: In an ideally segmented animal, all, , the segments are identical; such a condition is termed homonomous. No such animal exists, anal segment necessarily differ from the others,, , since one or more head segments and the, In most segmented animals, however, the, , but the polychaete annelids approach the ideal., , segmentation is heteronomous, i.e., has undergone local alterations through loss or fusion of, segments, emphasis of certain organs in some segments, loss of organs from others, migrations, of organs to other than the original segments, etc. Such alterations commonly begin anteriorly, and proceed posteriorly so that the posterior region of the animal usually retains the more, primitive condition. Commonly the head of segmented animals represents several fused, segments with emphasis on nervous structures and loss of representatives of other systems,, Primitively the number of segments is large and indefinite and becomes limited to a definite,, relatively small number in forms with a high degree of heteronomous segmentation., , 2. External and Internal Metamerism: Metamerism is conspicuously visible in most, annelids, both externally as well as internally. The common earthworm is a good illustration of {, both external and internal metamerism; the body consists of a great number of similar segments, andall the body organs, such as musculature, blood vessels, nerves, ganglia, excretory organs and, gonads, etc., are repeated segmentally. Even the coelom is divided into segmental compartments, by the intersegmental transverse mesenteries, called the septa. Only the digestive tract remains, unaffected, but it also extends through every segment. Among acanthocephalans, rotifers, and., some other “aschelminth” groups, external ring like formations, called annulations, occur in, the covering tissues, sometimes so marked as to suggest segmentations; these formations prove, to be only superficial, however, and are not indicative of true segmentation. In arthropods, too, metamerism is chiefly extemal, while man and other vertebrates show an internal metamerism, of body muscles, nerves, certain blood vessels, vertebrae and ribs. ae, , 3. Complete and Incomplete Metamerism: In annelid worms, metamerism is complete, ,, affecting practically all the systems. The metameres are essentially alike or homonomous, each, having segmental blood vessels, nerves, nephridia and coelomoducts. This is the primitive, condition. On the other hand, arthropods and vertebrates show incomplete metamerism. They:, are heteronomous animals, since division of labor has resulted in dissimilarity of the metameresof different regions of the body. Incomplete metamerism should not however be confused wi, the repetition of single organs, such as shell plates or gills, in certain unsegmented acirital, like the molluscs. Also, metamerism is more complete or uniform throughout the body of a, , , , Scanned with CamScanner
 Learn better on this topic
Learn better on this topic